Antibiotics possess potent antibacterial effects and are widely used in the prevention and treatment of human and animal diseases. Their antibacterial mechanisms primarily encompass hindering the synthesis of bacterial cell walls, altering the permeability of cell membranes, inhibiting bacterial ribosomes, and affecting the activity of RNA or other related enzymes. Natural antibiotics have been clinically used for a relatively long time, and their industrial production systems are becoming increasingly sophisticated, currently occupying a significant market share. Natural antibiotics can be classified into tetracyclines, macrolides, β-lactams, aminoglycosides, and other categories.
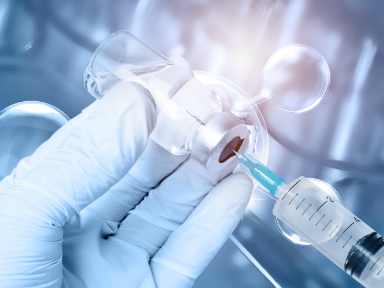
1. Tetracycline Antibiotics
Tetracycline antibiotics are a class of broad-spectrum antibiotics produced by actinomycetes. This type of antibiotic is primarily used in the treatment of infectious diseases caused by Gram-positive and Gram-negative bacteria, intracellular mycoplasmas, chlamydiae, and rickettsiae.
Ribosomes are the sites of protein synthesis in all living cells, composed of proteins and RNA. Numerous studies have shown that tetracycline can bind to the RNA component of bacterial ribosomes to exert its antibacterial activity. In other words, the antibacterial mechanism of tetracycline antibiotics involves their preferential binding to bacterial ribosomes and interacting with the highly conserved 16S rRNA target in the 30S ribosomal subunit to form a reversible complex. During the elongation process, this complex spatially interferes with the binding of charged aminoacyl-tRNA to the mRNA-ribosome complex, preventing new amino acids from entering the nascent peptide chain and inhibiting the synthesis of proteins essential for bacterial growth and survival, thereby achieving an antibacterial effect.
Resistance Mechanisms
Long-term and extensive use of tetracycline drugs can lead to significant bacterial resistance to this class of drugs. To date, 59 tetracycline resistance genes have been reported. They mediate resistance to tetracycline drugs through two distinct mechanisms. The first is through active efflux of tetracycline by efflux pumps. The most common tetracycline-specific efflux pumps are members of the major facilitator superfamily of transporters. The latest statistics indicate that there are 30 different tetracycline-specific efflux pumps in bacteria. Efflux pump genes encode membrane-associated efflux proteins that actively pump drugs out of the cell, resulting in a decrease in intracellular drug concentration, reduced binding of ribosomes to tetracycline drugs, and thus the development of resistance. The second mechanism involves the action of bacterial ribosomal protection proteins (RPPs). Tetracycline ribosomal protection proteins (RPPs), first described in Campylobacter jejuni and Streptococcus, are GTPases with significant sequence and structural similarity to elongation factors EF-G and EF-Tu. Binding of RPPs to ribosomes can reverse the distorted ribosomal structure, causing a conformational change in the ribosome that directly interferes with the stacking interaction between the tetracycline D-ring and the 16S rRNA base C1054. This prevents tetracycline drugs from binding and dissociates them from the 30S subunit binding site, thereby protecting the ribosome.
2.Aminoglycoside Antibiotics
Aminoglycosides are natural or semi-synthetic antibiotics extracted from actinomycetes. Since the first discovery of streptomycin, the first aminoglycoside antibiotic, in 1943, they have been widely used to treat infections caused by both Gram-negative and Gram-positive bacteria due to their excellent antibacterial activity and broad antibacterial spectrum.
Aminoglycoside antibiotics belong to the category of bacteriostatic drugs that act during the stationary phase of bacterial growth and can be used in combination with bacteriostatic drugs during the reproductive phase (β-lactam antibiotics) for synergistic antibacterial effects. The primary site of action of aminoglycoside antibiotics is also the ribosome, where they inhibit bacterial protein synthesis at the initiation, elongation, and termination stages. During the initiation of polypeptide chain synthesis in prokaryotes, aminoglycoside antibiotics bind to the 30S subunit, interfering with the assembly of the 30S initiation complex. During the elongation process of the polypeptide chain, aminoglycosides bind to the A site of the 30S subunit, causing misreading of the genetic code and leading to the synthesis of abnormal proteins. At the termination stage of polypeptide chain synthesis, aminoglycoside antibiotics prevent the binding of termination codons to ribosomes, further preventing the release of synthesized peptide chains and inhibiting the dissociation of 70S ribosomes, thereby blocking the protein synthesis cycle.
Resistance Mechanisms
Aminoglycoside resistance manifests in various forms, including inactivation of aminoglycoside antibiotics through enzymatic modification, alterations in target sites, and decreased membrane permeability.
①Changes in cell membrane permeability can lead to reduced drug uptake and accumulation in bacteria, resulting in bacterial resistance. The outer membrane of Gram-negative bacteria serves as a barrier to both hydrophobic and hydrophilic substances. To overcome this permeability barrier, these microorganisms have evolved porin proteins, which act as non-specific entry and exit points for antibiotics and other small organic chemicals. When the number of these porin proteins significantly decreases, bacteria develop resistance.
②Bacteria can produce aminoglycoside-modifying enzymes (AMEs) that inactivate aminoglycoside antibiotics, leading to antibiotic resistance. There are three types of AMEs: aminoglycoside nucleotidyltransferases (ANTs), aminoglycoside acetyltransferases (AACs), and aminoglycoside phosphotransferases (APHs). Enzymatic modification and inactivation are the most prevalent resistance mechanisms employed by bacteria against aminoglycoside antibiotics. AMEs covalently bind to functional groups such as -OH or -NH2, interfering with the binding of antibiotics to the A-site in 16S rRNA, which results in reduced affinity between antibiotics and ribosyl-tRNA, thereby inactivating the antibiotics.
③Ribosomes are the target sites of aminoglycoside antibiotics, and rRNA plays a crucial role in the binding of antibiotics to ribosomes. Alterations in the target site prevent antibiotics from binding to ribosomes. For instance, plasmid-mediated 16S rRNA methyltransferases (RMTs) modify specific rRNA nucleotide residues to prevent aminoglycosides from effectively binding to their sites, conferring high-level resistance to these drugs in bacteria.
3. Macrolide Antibiotics
Macrolide antibiotics are characterized by their typical structure containing a macrolide ring in their core. They are primarily effective against Gram-positive bacteria and have limited activity against Gram-negative bacteria. Their activity against eukaryotic organisms is low due to their weak affinity for eukaryotic ribosomes. The mechanism of action of macrolide antibiotics involves their irreversible binding to the 50S subunit of bacterial ribosomes, occupying a position within the nascent peptide exit tunnel (NPET) near the peptidyl transferase center. This binding inhibits protein synthesis.
Upon binding to the NPET, macrolides interfere with the peptide elongation phase by causing the dissociation of peptidyl-tRNA from the ribosome. This prevents ribosomes bound to macrolides from incorporating specific amino acid sequences into nascent proteins, thereby blocking the transpeptidation reaction and mRNA translocation processes. As a result, the elongation of the peptide chain is hindered, inhibiting protein synthesis and ultimately exerting a bacteriostatic effect.
Resistance Mechanisms of Macrolide Antibiotics
Currently, the resistance mechanisms of macrolide antibiotics primarily encompass two categories. Firstly, ribosome modification involves methyltransferases encoded by erythromycin-resistant methyltransferase genes, which catalyze the monomethylation or dimethylation of the N6 position in nucleotide A2058. This nucleotide interacts specifically with the sugar moiety at the C5 position of the macrolide ring. Methylation disrupts the formation of hydrogen bonds, leading to a significant decrease in the affinity between macrolides and the 50S subunit of ribosomes, thereby giving rise to resistant strains. Apart from rRNA methylation, rRNA mutations can also confer resistance. Mutations at A2058 alter the ribosomal target site, preventing the binding of macrolides.
Secondly, the intracellular concentration of macrolides can be reduced, enabling bacteria to evade the effects of these drugs. One way to achieve this is through the use of efflux pumps. The Mef and Msr subfamilies, closely related to macrolides, encode on plasmids and belong to the MSF and ATP-binding cassette (ABC) transporter families, respectively. The Mef pump functions as an antiporter, exchanging protons for bound macrolides. Instead of utilizing ATP as an energy source to pump antibiotics out of the cell, Mef proteins employ secondary active transport, where ATP energy is not directly used for transmembrane translocation of macrolides. Proteins of this subfamily are significant determinants of macrolide resistance, with Mef(A) and Mef(E) being the most common variants, both capable of conferring resistance to 14- and 15-membered macrolides. The Msr protein, on the other hand, provides ribosomal protection by binding to macrolide drugs.
4. β-Lactam Antibiotics
β-Lactam antibiotics represent the most widely used class of antibiotics available today. They all contain a four-membered β-lactam ring, often accompanied by a fused five- or six-membered ring. Based on the differences in their fused ring structures, β-lactam antibiotics are further categorized into penicillins, cephalosporins, and atypical β-lactams. Despite their structural variations, β-lactam antibiotics share a common mechanism of action. This mechanism primarily involves the inhibition of the transpeptidase activity of enzymes, which subsequently disrupts the synthesis of bacterial cell walls, ultimately leading to bacterial death.
Peptidoglycan, the primary component of bacterial cell walls, forms a net-like polymer that completely envelops the cell. It serves as the sole protective mechanism for Gram-positive bacteria against internal osmotic pressure, enabling them to survive in hypotonic environments. Recent research has revealed that bacterial cell membranes contain several specialized protein molecules that can form relatively stable complexes with penicillin, known as penicillin-binding proteins (PBPs). These PBPs represent the primary targets of β-lactam antibiotics. They are essential enzymes for the synthesis of peptidoglycan, a crucial component of the cell wall.
The structures of penicillin and cephalosporins resemble the terminal D-alanyl-D-alanine moiety of peptidoglycan, enabling them to competitively bind to the active centers of enzymes via covalent bonds. This binding inhibits the crosslinking reactions catalyzed by mucopeptide transpeptidases, severely disrupting the formation of bacterial cell walls. Consequently, this leads to bacterial lysis through a process known as osmolysis, ultimately resulting in bacterial death. Since mammalian cells lack cell walls, they are unaffected by this process, conferring a high degree of selectivity to the antibacterial action of these antibiotics.
Bacteria possess numerous intricate mechanisms that confer resistance to antibiotics. In the case of β-lactam antibiotics, bacteria develop resistance through three primary mechanisms, similar to most antimicrobials. These mechanisms are essential for Gram-positive bacteria to resist β-lactam antibiotics.
①Production of β-Lactamases: Bacteria produce β-lactamases that can covalently bind to the carbonyl moiety of antibiotics, destroying their cyclic structure. This results in the degradation of β-lactam antibiotics before they can reach their targets. Additionally, β-lactamases can rapidly and tightly bind to β-lactam antibiotics non-hydrolytically, preventing them from accessing their target sites and exerting their effects.
②Alteration of PBPs: PBPs are the targets of β-lactam drugs. When mutations occur in PBPs, the affinity between β-lactam antibiotics and their intended PBP targets is lost. Consequently, the drugs fail to bind to their action sites, thereby preventing them from exerting their therapeutic effects and leading to bacterial resistance.
③Changes in Outer Membrane Permeability: Modifications in cell membrane permeability or increased efflux pump activity can prevent antibiotics from entering bacterial cells. This reduction in antibiotic influx decreases the binding of drugs to their targets within the bacterial cells, subsequently lowering the active concentration of antibiotics within the cells. This mechanism also contributes to bacterial resistance.
References
[1] Feng Kai, Xin Jie, Tian Jun, Chang Honghong, Gao Wenchao. Research Progress on the Structure and Structure-Activity Relationships of Natural Antibiotics. Chinese Journal of Antibiotics, 2021, 46(09): 809-820. DOI: 10.13461/j.cnki.cja.007125.
[2] Tang Yuqing, Ye Qian, Zheng Weiyi. Current Status and Progress in the Research of Antibiotic Drugs. Foreign Medical Sciences (Antibiotics Section), 2019, 40(04): 295-301. DOI: 10.13461/j.cnki.wna.005229.
About the Author
Xiaonisha, a food technology professional holding a Master's degree in Food Science, is currently employed at a prominent domestic pharmaceutical research and development company. Her primary focus lies in the development and research of nutritional foods, where she contributes her expertise and passion to create innovative products.



 ALL
ALL Pharma in China
Pharma in China Pharma Experts
Pharma Experts Market News
Market News Products Guide
Products Guide Brand Story
Brand Story





























 Pharma Sources Insight July 2025
Pharma Sources Insight July 2025








